
medRxiv | Virlaza™ Inhibits Sars-COV-2-induced Inflammatory Response of Bronchial Epithelial Cells and Pulmonary Fibroblast
Coronavirus disease 2019 (COVID-19), which is currently a global public health emergency and beyond vaccines as a prophylactic treatment, no specific and effective therapeutical treatments are available. COVID-19 induces a massive release of proinflammatory cytokines, which drives COVID-19 progression, severity, and mortality. In addition, bronchial epithelial cells are the first pulmonary cells activated by coronavirus-2 (SARS-Cov-2) leading to massive cytokine release, which can hyperactivate lung fibroblasts, resulting in pulmonary fibrosis, a phenomenon observed even in moderate COVID-19 survivors. This in vitro study tested the hypothesis that Virlaza™, a herbal medicine, could inhibit the hyperactivation of human bronchial epithelial cells (BEAS-2B) and pulmonary fibroblasts (MRC-5) induced by SARS-Cov-2. BEAS-2B (5×104/mL/well) and MRC-5 (5×104/mL/well) cells were co-cultivated with 1ml of blood of a Sars-Cov-2 infected patient for 4 hours and Virlaza™ (1ug/mL) was added in the first minute of the co-culture. After 4 hours, the cells were recovered and used for analysis of cytotoxicity by MTT and for mRNA expression of P2X7 receptor E iNOS. The supernatant was used to measure ATP and cytokines. Sars-Cov-2 incubation resulted in increased release of ATP, IL-1beta, IL-6, IL-8, and TNF-alpha by BEAS-2B and MRC-5 cells (p<0.001). Treatment with Virlaza™ resulted in reduction of ATP, IL-1beta, IL-6, IL-8, and TNF-alpha release (p<0.001). In addition, Sars-Cov-2 incubation resulted in increased expression of P2X7 receptor and iNOS (p<0.001), which has been reversed by Virlaza™ (p<0.001). In conclusion, Virlaza™ presents important anti-inflammatory effects in the context of Sars-Cov-2 infection.
Abstract
Introduction
The quick spread of Coronavirus disease (COVID-19) resulted in a pandemic in the whole world, reaching more than 100 million cases and 2 million deaths due to the severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) [1]. The progression, severity, and mortality of COVID-19 differ from no, mild, or moderate symptoms in most of the patients, while up to 15% develop the severe and critical form of the disease with high mortality rates [2]. Among the factors underlying the development of the severe and critical forms of COVID-19, advanced age [3], comorbidities [3], hyperactivation of the immune system [3,4] are proven to have a central role, hyperactivation of the immune system results in the cytokine storm, which is characterized by the synthesis and release of high levels of proinflammatory cytokines [3,4]. In this way, cytokine storm plays a major role in Covid-19 pathogenesis [3,4].
In this context, bronchial epithelial cells have been described not only as a mechanical barrier for the respiratory tract, but also as an important cell layer centrally involved in the innate immune response, which actively synthetize and release a plethora of inflammatory and fibrotic mediators [5], capable to activate fibroblasts inducing fibrosis. In addition, activation of the purinergic receptor P2X7 in the airway epithelium has been described as a key receptor involved in the inflammatory [6] and fibrotic [7] responses of the lungs to a variety of stimulus, including SARS-CoV-2 [8]. Furthermore, P2X7 is also involved in pulmonary fibroblast proliferation and activation, driven the pulmonary fibrotic response as well [7].
Activation of P2X7 receptor may be a trigger by increased levels of nitric oxide (NO) mediated by increased expression and activation of inducible nitric oxide synthase (iNOS) [9]. Indeed, increased iNOS expression drives an increased inflammatory and fibrotic responses of the lungs, such as in asthma [10], pulmonary fibrosis [11], and bacterial infections [12, 13].
Thus, this study tested the hypothesis that Virlaza™ could inhibit bronchial epithelial cells and lung fibroblast activation in vitro, involving iNOS and P2X7 signaling.
In this context, bronchial epithelial cells have been described not only as a mechanical barrier for the respiratory tract, but also as an important cell layer centrally involved in the innate immune response, which actively synthetize and release a plethora of inflammatory and fibrotic mediators [5], capable to activate fibroblasts inducing fibrosis. In addition, activation of the purinergic receptor P2X7 in the airway epithelium has been described as a key receptor involved in the inflammatory [6] and fibrotic [7] responses of the lungs to a variety of stimulus, including SARS-CoV-2 [8]. Furthermore, P2X7 is also involved in pulmonary fibroblast proliferation and activation, driven the pulmonary fibrotic response as well [7].
Activation of P2X7 receptor may be a trigger by increased levels of nitric oxide (NO) mediated by increased expression and activation of inducible nitric oxide synthase (iNOS) [9]. Indeed, increased iNOS expression drives an increased inflammatory and fibrotic responses of the lungs, such as in asthma [10], pulmonary fibrosis [11], and bacterial infections [12, 13].
Thus, this study tested the hypothesis that Virlaza™ could inhibit bronchial epithelial cells and lung fibroblast activation in vitro, involving iNOS and P2X7 signaling.
Materials and Methods
This study and all experimental procedures were analyzed and approved by ethical committee of School of Medicine of Anhembi Morumbi University (4.637.625 in 08 of April of 2021) and were carried out in accordance with Declaration of Helsinki for ethical principles for medical research involving human subjects.
Human Cell Co-Culture Study
Virlaza™ [Clove glycerin extract (1:5); Eucalyptus Glycerin Extract (1:5); Basil Glycerin Extract (1:5); Sage Glycerin Extract (1:5); Maritime pine Glycerin extract (1:5); Clove Tincture (ethyl alcohol 99.8%) (1:5); Eucalyptus Tincture (1:5); Maritime pine Tincture (ethyl alcohol 99.8%) (1:5); Basilica Tincture (ethyl alcohol 99.8%); Sage Tincture (ethyl alcohol 99.8%) (1:5); Selenite (0.0051%), and Zinc (0.32%).
Human bronchial epithelial cell line (BEAS-2B) and human lung fibroblast cell line (MRC-5), which are commercially available of immortalized cell lines, were co-cultured with whole blood from an infected patient with Sars-Cov-2. BEAS-2B and MRC-5 cells were obtained from Rio de Janeiro Cell Bank and co-cultured in DMEM high glucose medium (Sigma Chemical Co., St. Louis, MO, USA) supplemented with 10% fetal calf serum in 500ul of whole blood of Sars-Cov-2 infected male (35-40) years old patient (original Wuhan Sars-Cov-2), presenting an estimated high viral load based on the cycle threshold (CT = 15). Both BEAS-2B and MRC-5 cells were co-cultured at a concentration of 5×104/mL/well in 48 wells plate, at a humidified atmosphere in a CO2 incubator (5% CO2, 37°C) [14]. The blood was collected immediately after hospitalization, prior any antibiotic and corticosteroid administration. The written consent inform was obtained from the patient volunteer for this study. The cells (BEAS-2B or MRC-5) were incubated at the same time with Virlaza™ (1ug/mL) and with 500ul of whole blood of Sars-Cov-2 infected patients and the cells and supernatant was obtained 4 hours after stimulation. The cells and supernatant were recovered, centrifuged at 900g for 5 minutes at 4°C, and then the supernatant immediately used for ATP measurement and later for cytokine measurement. The cells were washed using phosphate-buffered saline (PBS) and then subjected to RT-PCR protocol.
Human bronchial epithelial cell line (BEAS-2B) and human lung fibroblast cell line (MRC-5), which are commercially available of immortalized cell lines, were co-cultured with whole blood from an infected patient with Sars-Cov-2. BEAS-2B and MRC-5 cells were obtained from Rio de Janeiro Cell Bank and co-cultured in DMEM high glucose medium (Sigma Chemical Co., St. Louis, MO, USA) supplemented with 10% fetal calf serum in 500ul of whole blood of Sars-Cov-2 infected male (35-40) years old patient (original Wuhan Sars-Cov-2), presenting an estimated high viral load based on the cycle threshold (CT = 15). Both BEAS-2B and MRC-5 cells were co-cultured at a concentration of 5×104/mL/well in 48 wells plate, at a humidified atmosphere in a CO2 incubator (5% CO2, 37°C) [14]. The blood was collected immediately after hospitalization, prior any antibiotic and corticosteroid administration. The written consent inform was obtained from the patient volunteer for this study. The cells (BEAS-2B or MRC-5) were incubated at the same time with Virlaza™ (1ug/mL) and with 500ul of whole blood of Sars-Cov-2 infected patients and the cells and supernatant was obtained 4 hours after stimulation. The cells and supernatant were recovered, centrifuged at 900g for 5 minutes at 4°C, and then the supernatant immediately used for ATP measurement and later for cytokine measurement. The cells were washed using phosphate-buffered saline (PBS) and then subjected to RT-PCR protocol.
Cytotoxicity assay by MTT assay
To assess the Virlaza™ cytotoxicity through cell viability, MTT assay was carried out. Briefly, 5×104 viable BEAS-2B and MRC-5 cells were placed into clear 96-well flat-bottom plates (Corning USA) in DMEM high glucose medium supplemented with 10% fetal calf serum and immediately after different concentrations (0.1ug/mL; 1ug/mL; 10ug/mL; 100ug/mL; and 1000ug/mL) of Virlaza™. Following 24h after incubation in a humidified atmosphere of a CO2 incubator (5% CO2, 37°C), 10μL/well of MTT (5mg/mL) was added to the cells (both in the control and Virlaza™), which was incubated for 4h. After this time, 100μL of 10% sodium dodecyl sulfate (SDS) solution in deionized water was added to the cells and incubated overnight [15, 16]. The absorbance was measured at 595nm in a benchtop multimode reader SpectraMax i3 (Molecular Devices, San Jose, CA, USA).
Adenosine Triphosphate (ATP) Measurement
The ATP concentration was determined in the cell culture supernatant immediately after its collection by using the ATPlite Luminescence Assay System (Perkin Elmer, Waltham, MA, USA), according to the manufacturer’s instructions. The results were expressed in nmol/mL [17].
Cytokines Measurement
The levels of interleukin (IL)-1beta (DY201), IL-1RA (DY280), IL-6 (DY206), IL-8 (DY208) and TNF-alpha (DY210) were measured in the cell culture supernatant by using DuoSet ELISA kit (R&D Systems; Minneapolis, MN, USA) according to the manufacturer’s recommendations using the SpectraMax i3 multiplate reader (Molecular Devices, CA, USA). The results were expressed as pg/mL [14-17].
Reverse transcriptase-polymerase chain reaction (RT-PCR)
Total RNA isolation from cell pellets was used RNeasy mini kits (Qiagen, Hilden, Germany). Reverse transcription was used Stratascript reverse transcriptase (Stratagene, CA, USA) and random primers (Invitrogen, Germany). Quantitative PCR used Taqman Universal PCR Mastermix (Applied Biosystems, USA) and preformulated primers and probe mixes (Assay on Demand, Applied Biosystems, USA). PCR conditions were 2 min at 50°C, 10 min at 95°C, followed by 45 cycles of 15 s at 95°C and 60°C for 1 min using the CFX96 thermal cycler (Bio rad, Hercules CA, USA). PCR amplification of the housekeeping gene encoding Glyceraldehyde-3-phosphate dehydrogenase (GADPH) was performed during each run for each sample to allow normalization between samples. β-actin was used as a control and correction factor for the expression of the P2X7 receptor, for which the sequences of primers are described as follows: β-actin - forward (5′GTGGGCCGCTCTAGGCACCA3′) and reverse primers (5′CTCTTTGATGTCACGCACGATTTC3′ 540 bp) [15] and P2X7 - forward, 5′ AGATCGTGGAGAATGGAGTG 3′, and (5′ TTCTCGTGGTGTAGTTGTGG 3′) reverse primers [15]. For iNOS – forward 5′-CTATCAGGAAGAAATGCAGGAGAT-3′, and Reverse 5′-GAGCACGCTGAGTACCTCATT-3′ primers [16].
Statistical Analysis
The software Graph Pad Prism 5.0 was used to perform the statistical analysis and build de graphs. The results were expressed as mean ± standard□error□of□the mean□(SEM) from at least three independent experiments. One-way analysis of variance (ANOVA) was used for multiple comparisons, followed by the Bonferroni post hoc test for comparison among groups. A p-value <0.05 was considered significant.
Results
Figure 1 shows that Sars-Cov-2 activated BEAS-2B and MRC-5 cells, resulting in increased release of ATP by BEAS-2B (Figure 1, A; p<0.001) and by MRC-5 (Figure 1 B; p<0.001) cells, which has been significantly attenuated by Virlaza™, both in BEAS-2B (Figure 1 A; p<0.001) and by MRC-5 (Figure 1 B; p<0.001) cells.
Effects of Virlaza™ on ATP Release in BEAS-2B and MRC-5 Cells
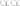
{kind=link}
{kind=link}
Figure 1
Effects of Virlaza™ on ATP Release in BEAS-2B and MRC-5 Cells
Adenosine triphosphate (ATP) levels in the supernatant of the cells. Figure 1 A correspond to experiments done in BEAS-2B cells. Figure 1 A correspond to experiments done in MRC-5 cells. *** p<0.0001. Co = Control; Vir = Treated with 10ug/mL of Virlaza™; SarsCov2 = Stimulated with blood infected with Sars-Cov-2; SarsCov2+Vir = Stimulated with blood infected with Sars-Cov- 2 + 10ug/mL of Virlaza™.
Effects of Virlaza™ on ATP Release in BEAS-2B and MRC-5 Cells
Adenosine triphosphate (ATP) levels in the supernatant of the cells. Figure 1 A correspond to experiments done in BEAS-2B cells. Figure 1 A correspond to experiments done in MRC-5 cells. *** p<0.0001. Co = Control; Vir = Treated with 10ug/mL of Virlaza™; SarsCov2 = Stimulated with blood infected with Sars-Cov-2; SarsCov2+Vir = Stimulated with blood infected with Sars-Cov- 2 + 10ug/mL of Virlaza™.
Effects of Virlaza™ on Bronchial Epithelial Cells (BEAS-2B) Activation
Figure 2 shows that Sars-Cov-2 activation resulted in increased synthesis and release of IL-1beta (Figure 2, A; p<0.001), IL-6 (Figure 2 C; p<0.001), IL-8 (Figure 2 D; p<0.001), and TNF-alpha (Figure 2 E; p<0.001), while Virlaza™ resulted in a significant reduction of the synthesis and release of all these pro-inflammatory cytokines (p<0.001). On the other hand, on Sars-Cov-2 stimulated cells, Virlaza™ resulted in a significant increase in the synthesis and release of IL-1RA (Figure 2 B; p<0.001), a potent anti-inflammatory cytokine.
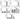
Figure 2
Effects of Virlaza™ on Bronchial Epithelial Cells (BEAS-2B) Activation
Cytokines levels in the supernatant of BEAS-2B cells. *p<0.05; **p<0.01; *** p<0.0001. Co = Control; Vir = Treated with 10ug/mL of Virlaza™; SarsCov2 = Stimulated with blood infected with Sars-Cov-2; SarsCov2+Vir = Stimulated with blood infected with Sars-Cov-2 + 10ug/mL of Virlaza™.
Effects of Virlaza™ on Bronchial Epithelial Cells (BEAS-2B) Activation
Cytokines levels in the supernatant of BEAS-2B cells. *p<0.05; **p<0.01; *** p<0.0001. Co = Control; Vir = Treated with 10ug/mL of Virlaza™; SarsCov2 = Stimulated with blood infected with Sars-Cov-2; SarsCov2+Vir = Stimulated with blood infected with Sars-Cov-2 + 10ug/mL of Virlaza™.
Effects of Virlaza™ on Pulmonary Fibroblasts (MRC-5) Activation
Figure 3 shows that Sars-Cov-2 activation resulted in increased synthesis and release of IL-1beta (Figure 3 A; p<0.001), IL-6 (Figure 3 C; p<0.001), and TNF-alpha (Figure 3 E; p<0.001), but not of IL-8 (Figure 3 D; p>0;05), while Virlaza™ resulted in a significant reduction of the synthesis and release of all these pro-inflammatory cytokines (p<0.001). On the other hand, in Sars-Cov-2 stimulated cells, Virlaza™ resulted in a significant increase in the synthesis and release of IL-1RA (Figure 3 B; p<0.001), a potent anti-inflammatory cytokine.
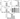
Figure 3
Effects of Virlaza™ on Pulmonary Fibroblasts (MRC-5) Activation
Cytokines levels in the supernatant of MRC-5 cells. *** p<0.0001. Co = Control; Vir = Treated with 10ug/mL of Virlaza™; SarsCov2 = Stimulated with blood infected with Sars-Cov-2; SarsCov2+Vir = Stimulated with blood infected with Sars-Cov-2 + 10ug/mL of Virlaza™.
Effects of Virlaza™ on Pulmonary Fibroblasts (MRC-5) Activation
Cytokines levels in the supernatant of MRC-5 cells. *** p<0.0001. Co = Control; Vir = Treated with 10ug/mL of Virlaza™; SarsCov2 = Stimulated with blood infected with Sars-Cov-2; SarsCov2+Vir = Stimulated with blood infected with Sars-Cov-2 + 10ug/mL of Virlaza™.
Effects of Virlaza™ on P2X7 receptor expression in BEAS-2B and MRC-5 Cells
Figure 4 shows that Sars-Cov-2 activation resulted in increased expression of P2X7 receptor in BEAS-2B (Figure 4 A; p<0.001), and in MRC-5 (Figure 4 B; p<0.001), while Virlaza™ resulted in a significant reduction of the expression of P2X7 receptor in BEAS-2B (Figure 4 A; p<0.001), and in MRC-5 (Figure 4 B; p<0.001) cells.
{kind=link}
{kind=link}
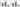
Figure 4
Effects of Virlaza™ on P2X7 receptor expression in BEAS-2B and MRC-5 Cells
P2X7 Receptor mRNA expression in BEAS-2B and MRC-5 cells. **p<0.01; *** p<0.0001. Co = Control; Vir = Treated with 10ug/mL of Virlaza™; SarsCov2 = Stimulated with blood infected with Sars-Cov-2; SarsCov2+Vir = Stimulated with blood infected with Sars-Cov-2 + 10ug/mL of Virlaza™.
Effects of Virlaza™ on iNOS mRNA Expression in BEAS-2B and MRC-5 Cells
Figure 5 shows that Sars-Cov-2 activation resulted in increased expression of iNOS in BEAS-2B (Figure 5 A; p<0.001), and in MRC-5 (Figure 5 B; p<0.001), while Virlaza™ resulted in a significant reduction of the expression of iNOS in BEAS-2B (Figure 5 A; p<0.001), and in MRC-5 (Figure 5 B; p<0.001) cells.
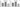
Figure 5
Effects of Virlaza™ on iNOS mRNA Expression in BEAS-2B and MRC-5 Cells
P2X7 Receptor mRNA expression in BEAS-2B and MRC-5 cells. *p<0.05; **p<0.01; *** p<0.0001. Co = Control; Vir = Treated with 10ug/mL of Virlaza™; SarsCov2 = Stimulated with blood infected with Sars-Cov-2; SarsCov2+Vir = Stimulated with blood infected with Sars-Cov-2 + 10ug/mL of Virlaza™.
Effects of Virlaza™ on iNOS mRNA Expression in BEAS-2B and MRC-5 Cells
P2X7 Receptor mRNA expression in BEAS-2B and MRC-5 cells. *p<0.05; **p<0.01; *** p<0.0001. Co = Control; Vir = Treated with 10ug/mL of Virlaza™; SarsCov2 = Stimulated with blood infected with Sars-Cov-2; SarsCov2+Vir = Stimulated with blood infected with Sars-Cov-2 + 10ug/mL of Virlaza™.
Effects of Virlaza™ on Cell Viability of BEAS-2B and MRC-5 Cells
Figure 6 shows the different concentrations of Virlaza™ on cell toxicity to determine the IC50 value. Figures 6 A (BEAS-2B cells) and 6 B (MRC-5 cells) show that 10 μg/mL was reached, was the dose corresponding to IC50, and was chosen was the study dose.
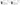
Figure 6
Effects of Virlaza™ on Cell Viability of BEAS-2B and MRC-5 Cells
Cell viability (%) measured by MMT assay in BEAS-2B and MRC-5 cells stimulated with growing doses of Virlaza™ (0.1ug/mL, 1ug/mL, 10ug/mL, 100ug/mL, 1000ug/mL).
Effects of Virlaza™ on Cell Viability of BEAS-2B and MRC-5 Cells
Cell viability (%) measured by MMT assay in BEAS-2B and MRC-5 cells stimulated with growing doses of Virlaza™ (0.1ug/mL, 1ug/mL, 10ug/mL, 100ug/mL, 1000ug/mL).
Discussion
This study shows for the first time that Virlaza™ was able to block the cytokine storm caused by Sars-Cov-2-induced epithelial (BEAS-2B) and lung fibroblast (MRC-5) activation, involving dampening of exacerbated purinergic signaling and iNOS expression. In addition, the study demonstrated the safety of an active therapeutic dose Virlaza™ in the context of cellular toxicity.
Considering the lack of effective treatment for COVID-19, and the evidence that severely ill COVID-19 patients develop a cytokine storm, some studies are testing the hypothesis that the blockade of some proinflammatory cytokines, such as IL-6 [18] and IL-1beta [19] could be beneficial for severely ill COVID-19 patients. In addition, beyond massive infiltration by neutrophils and macrophages into the lungs, increased blood levels of IL-1β, IL-6, and TNF-alpha have been linked to higher severity and mortality in COVID-19 patients [18, 19]. In this context, bronchial epithelial cells are the main entrance door and the main target of different types of respiratory bacteria and viruses, harming the immune system by exacerbating release of pro-inflammatory cytokines and growth factors [20, 21]. In the present study, we demonstrated that bronchial epithelial cells (BEAS-2B) when stimulated with Sars-Cov-2, responded to increased release of IL-1beta, IL-6, IL-8 and TNF-alpha, which were abolished by treatment with Virlaza™, clearly demonstrating the efficacy of Virlaza™ blocking the epithelial hyperactivation in the context of Sars-Cov-2.
Importantly, the literature demonstrates the importance of increased expression of P2X7 receptor and consequent activation triggering proinflammatory and pro-fibrotic response in the context of respiratory diseases [4-7]. Such importance has been demonstrated in several cells of the hematopoietic system [6], but also in lung structural cells, such as in epithelial cells [4, 5] and lung fibroblasts [7]., the activation of P2X7 receptors is classically related to the initiation, maintenance, progression, and severity of inflammatory states triggering IL-1beta release [7-9]. Herein, this study demonstrated that Virlaza™ inhibited IL-1beta release not only by epithelial cells (BEAS-2B) but also by lung fibroblasts (MRC-5). Particularly about fibroblasts, the inhibitory effect of Virlaza™ inhibiting the release of IL-1beta is of particular importance, since IL-1beta directly stimulates collagen synthesis and proliferation in fibroblasts [22, 23]. Therefore, it is plausible to hypothesize that Virlaza™ could also have a potential effect to inhibit lung fibrosis, which is commonly observed in COVID-19 survivors [24]. Such observed effects of Virlaza™ on IL-1beta release can be supported by the additional results of the present study, which demonstrated that Virlaza™ inhibited ATP release and accumulation, resulting in decreased expression of P2X7 receptor in BEAS-2B and in MRC-5 cells. Therefore, further clinical trial investigating the effects of Virlaza™ in COVID-19 survivors were guaranteed.
Beyond P2X7 receptor, inducible nitric oxide synthase (iNOS), is though as a key enzyme involved in all aspects of the pathophysiological process of pulmonary infections with diverse etiologies, including for the Sars-Cov-2 [25, 26]. In the context of COVID-19, intense and diffuse pulmonary inflammation results in endothelial dysfunction of the lung vasculature, uncoupling eNOS activity, lowering nitric oxide (NO) production, resulting in different pulmonary alterations and coagulopathy [25, 26]. On the contrary, viral infections trigger an increasing in iNOS activity, which may be acutely advantageous for host defense, once NO plays antiviral effects. However, sustained overproduction of NO mediates deleterious proinflammatory effects [25, 26]. In the present study, it was observed that Sars-Cov-2 increased iNOS expression in epithelial cells (BEAS-2B) and in lung fibroblasts (MRC-5 cells), which was dampened by Virlaza™. Interestingly, the literature demonstrates that NO synthesized specifically by the airway epithelium is vital to antiviral, inflammatory, and immune defense of the lungs [26]. However, again, the increased NO mediated by iNOS is related to an unresolved inflammatory process [27] as well as pro-fibrotic response [28]. iNOS overexpression is linked to the release of proinflammatory cytokines, such as IL-1beta, IL-6, IL-8, and TNF-alpha, particularly in the context of acute respiratory distress syndrome (ARDS), phenomena present in cases of severe COVID-19 [27]. In the present study, Sars-Cov-2 stimulation resulted in an increase expression of iNOS by epithelial cells (BEAS-2B) and in lung fibroblasts (MRC-5 cells) and increased release of IL-1beta, IL-6, IL-8, and TNF-alpha, Virlaza™ significantly reduced iNOS expression and pro-inflammatory cytokines release, reinforcing the anti-inflammatory and anti-fibrotic effects of Virlaza™. These findings concerning the iNOS inhibition by Virlaza™ can be strengthened once the literature already points out the possible therapeutic role of iNOS blockade for acute and long COVID-19 [29].
Beyond to inhibit the release of proinflammatory cytokines (IL-1beta, IL-6, IL-8, and TNF-alpha), Virlaza™ also induced the release of anti-inflammatory cytokine IL-1RA by epithelial cells (BEAS-2B) and by lung fibroblasts (MRC-5 cells). Functionally, interleukin-1 receptor antagonist (IL-1RA) inhibits the activities of interleukin 1 alpha (IL-1alpha) and interleukin 1 beta (IL-1beta), modulating a wide range of immune and inflammatory responses driven by interleukin 1 related cytokines [30]. Anakinra is a recombinant IL-1RA that has been tested for the treatment of COVID-19 patients, resulting in better clinical outcomes [31-33].
Therefore, the present study concludes that Virlaza™ was capable not only to inhibit the release of proinflammatory cytokines but also to stimulate the release of the anti-inflammatory cytokine IL-1RA, which has already been demonstrated to be a promising therapeutic option to prevent the development of severe forms of COVID-19 including respiratory failure, as well to treat the severe forms of COVID-19, improving the clinical outcomes and survival [31-33]. Thus, based on the findings of the present study, further clinical trials with Virlaza™ for the prevention and treatment of COVID-19 are guaranteed.
Considering the lack of effective treatment for COVID-19, and the evidence that severely ill COVID-19 patients develop a cytokine storm, some studies are testing the hypothesis that the blockade of some proinflammatory cytokines, such as IL-6 [18] and IL-1beta [19] could be beneficial for severely ill COVID-19 patients. In addition, beyond massive infiltration by neutrophils and macrophages into the lungs, increased blood levels of IL-1β, IL-6, and TNF-alpha have been linked to higher severity and mortality in COVID-19 patients [18, 19]. In this context, bronchial epithelial cells are the main entrance door and the main target of different types of respiratory bacteria and viruses, harming the immune system by exacerbating release of pro-inflammatory cytokines and growth factors [20, 21]. In the present study, we demonstrated that bronchial epithelial cells (BEAS-2B) when stimulated with Sars-Cov-2, responded to increased release of IL-1beta, IL-6, IL-8 and TNF-alpha, which were abolished by treatment with Virlaza™, clearly demonstrating the efficacy of Virlaza™ blocking the epithelial hyperactivation in the context of Sars-Cov-2.
Importantly, the literature demonstrates the importance of increased expression of P2X7 receptor and consequent activation triggering proinflammatory and pro-fibrotic response in the context of respiratory diseases [4-7]. Such importance has been demonstrated in several cells of the hematopoietic system [6], but also in lung structural cells, such as in epithelial cells [4, 5] and lung fibroblasts [7]., the activation of P2X7 receptors is classically related to the initiation, maintenance, progression, and severity of inflammatory states triggering IL-1beta release [7-9]. Herein, this study demonstrated that Virlaza™ inhibited IL-1beta release not only by epithelial cells (BEAS-2B) but also by lung fibroblasts (MRC-5). Particularly about fibroblasts, the inhibitory effect of Virlaza™ inhibiting the release of IL-1beta is of particular importance, since IL-1beta directly stimulates collagen synthesis and proliferation in fibroblasts [22, 23]. Therefore, it is plausible to hypothesize that Virlaza™ could also have a potential effect to inhibit lung fibrosis, which is commonly observed in COVID-19 survivors [24]. Such observed effects of Virlaza™ on IL-1beta release can be supported by the additional results of the present study, which demonstrated that Virlaza™ inhibited ATP release and accumulation, resulting in decreased expression of P2X7 receptor in BEAS-2B and in MRC-5 cells. Therefore, further clinical trial investigating the effects of Virlaza™ in COVID-19 survivors were guaranteed.
Beyond P2X7 receptor, inducible nitric oxide synthase (iNOS), is though as a key enzyme involved in all aspects of the pathophysiological process of pulmonary infections with diverse etiologies, including for the Sars-Cov-2 [25, 26]. In the context of COVID-19, intense and diffuse pulmonary inflammation results in endothelial dysfunction of the lung vasculature, uncoupling eNOS activity, lowering nitric oxide (NO) production, resulting in different pulmonary alterations and coagulopathy [25, 26]. On the contrary, viral infections trigger an increasing in iNOS activity, which may be acutely advantageous for host defense, once NO plays antiviral effects. However, sustained overproduction of NO mediates deleterious proinflammatory effects [25, 26]. In the present study, it was observed that Sars-Cov-2 increased iNOS expression in epithelial cells (BEAS-2B) and in lung fibroblasts (MRC-5 cells), which was dampened by Virlaza™. Interestingly, the literature demonstrates that NO synthesized specifically by the airway epithelium is vital to antiviral, inflammatory, and immune defense of the lungs [26]. However, again, the increased NO mediated by iNOS is related to an unresolved inflammatory process [27] as well as pro-fibrotic response [28]. iNOS overexpression is linked to the release of proinflammatory cytokines, such as IL-1beta, IL-6, IL-8, and TNF-alpha, particularly in the context of acute respiratory distress syndrome (ARDS), phenomena present in cases of severe COVID-19 [27]. In the present study, Sars-Cov-2 stimulation resulted in an increase expression of iNOS by epithelial cells (BEAS-2B) and in lung fibroblasts (MRC-5 cells) and increased release of IL-1beta, IL-6, IL-8, and TNF-alpha, Virlaza™ significantly reduced iNOS expression and pro-inflammatory cytokines release, reinforcing the anti-inflammatory and anti-fibrotic effects of Virlaza™. These findings concerning the iNOS inhibition by Virlaza™ can be strengthened once the literature already points out the possible therapeutic role of iNOS blockade for acute and long COVID-19 [29].
Beyond to inhibit the release of proinflammatory cytokines (IL-1beta, IL-6, IL-8, and TNF-alpha), Virlaza™ also induced the release of anti-inflammatory cytokine IL-1RA by epithelial cells (BEAS-2B) and by lung fibroblasts (MRC-5 cells). Functionally, interleukin-1 receptor antagonist (IL-1RA) inhibits the activities of interleukin 1 alpha (IL-1alpha) and interleukin 1 beta (IL-1beta), modulating a wide range of immune and inflammatory responses driven by interleukin 1 related cytokines [30]. Anakinra is a recombinant IL-1RA that has been tested for the treatment of COVID-19 patients, resulting in better clinical outcomes [31-33].
Therefore, the present study concludes that Virlaza™ was capable not only to inhibit the release of proinflammatory cytokines but also to stimulate the release of the anti-inflammatory cytokine IL-1RA, which has already been demonstrated to be a promising therapeutic option to prevent the development of severe forms of COVID-19 including respiratory failure, as well to treat the severe forms of COVID-19, improving the clinical outcomes and survival [31-33]. Thus, based on the findings of the present study, further clinical trials with Virlaza™ for the prevention and treatment of COVID-19 are guaranteed.
Data Availability
The raw data will be available upon a rasonable request.
References
1.Poor HD. Pulmonary Thrombosis and Thromboembolism in COVID-19. Chest. 2021 Jun 18:S0012-3692(21)01126-0. doi: 10.1016/j.chest.2021.06.016. Epub ahead of print. PMID: 34153340.
CrossRefPubMedGoogle Scholar
2.de Roquetaillade C, Mansouri S, Brumpt C, Neuwirth M, Voicu S, Le Dorze M, Fontaine C, Barthelemy R, Gayat E, Megarbane B, Mebazaa A, Chousterman BG. Comparison of Circulating Immune Cells Profiles and Kinetics Between Coronavirus Disease 2019 and Bacterial Sepsis. Crit Care Med. 2021 May 18. doi: 10.1097/CCM.0000000000005088. Epub ahead of print. PMID: 34001691.
CrossRefPubMedGoogle Scholar
3.Lange A, Lange J, Jaskuła E. Cytokine Overproduction and Immune System Dysregulation in alloHSCT and COVID-19 Patients. Front Immunol. 2021 Jun 2;12:658896. doi: 10.3389/fimmu.2021.658896. PMID: 34149697; PMCID: PMC8206782.
CrossRefPubMedGoogle Scholar
4.Chua RL, Lukassen S, Trump S, Hennig BP, Wendisch D, Pott F, Debnath O, Thürmann L, Kurth F, Völker MT, Kazmierski J, Timmermann B, Twardziok S, Schneider S, Machleidt F, Müller-Redetzky H, Maier M, Krannich A, Schmidt S, Balzer F, Liebig J, Loske J, Suttorp N, Eils J, Ishaque N, Liebert UG, von Kalle C, Hocke A, Witzenrath M, Goffinet C, Drosten C, Laudi S, Lehmann I, Conrad C, Sander LE, Eils R. COVID-19 severity correlates with airway epithelium-immune cell interactions identified by single-cell analysis. Nat Biotechnol. 2020 Aug;38(8):970–979. doi: 10.1038/s41587-020-0602-4. Epub 2020 Jun 26. PMID: 32591762.
CrossRefPubMedGoogle Scholar
5.Frey A, Lunding LP, Ehlers JC, Weckmann M, Zissler UM, Wegmann M. More Than Just a Barrier: The Immune Functions of the Airway Epithelium in Asthma Pathogenesis. Front Immunol. 2020 Apr 28;11:761. doi: 10.3389/fimmu.2020.00761. eCollection 2020.
CrossRefGoogle Scholar
6.Leyva-Grado VH, Ermler ME, Schotsaert M, Gonzalez MG, Gillespie V, Lim JK, GarcÍa-Sastre A. Contribution of the Purinergic Receptor P2X7 to Development of Lung Immunopathology during Influenza Virus Infection. mBio. 2017 Mar 28;8(2):e00229–17. doi: 10.1128/mBio.00229-17.
CrossRefGoogle Scholar
7.Riteau N, Gasse P, Fauconnier L, Gombault A, Couegnat M, Fick L, Kanellopoulos J, Quesniaux VF, Marchand-Adam S, Crestani B, Ryffel B, Couillin I. Extracellular ATP is a danger signal activating P2X7 receptor in lung inflammation and fibrosis. Am J Respir Crit Care Med. 2010 Sep 15;182(6):774–83. doi: 10.1164/rccm.201003-0359OC. Epub 2010.
CrossRefPubMedWeb of ScienceGoogle Scholar
8.Di Virgilio F, Tang Y, Sarti AC, Rossato M. A rationale for targeting the P2X7 receptor in Coronavirus disease 19. Br J Pharmacol. 2020 Nov;177(21):4990–4994. doi: 10.1111/bph.15138. Epub 2020 Jul 26.
CrossRefGoogle Scholar
9.Chiao CW, Tostes RC, Webb RC. P2X7 receptor activation amplifies lipopolysaccharide-induced vascular hyporeactivity via interleukin-1 beta release. J Pharmacol Exp Ther. 2008 Sep;326(3):864–70. doi: 10.1124/jpet.107.135350. Epub 2008 Jun 16.
Abstract/FREE Full TextGoogle Scholar
10.Sato Y, Chibana K, Horigane Y, Uchida N, Masawa M, Koike R, Nakamura Y, Watanabe T, Shiobara T, Arai R, Shimizu Y, Takemasa A, Ishii Y. Comparison of inducible nitric oxide synthase mRNA expression in different airway portions and association with nitric oxide parameters from patients with asthma. Clin Exp Allergy. 2019 May;49(5):582–590. doi: 10.1111/cea.13344. Epub 2019 Feb 15.
CrossRefGoogle Scholar
11.Venosa A, Gow JG, Taylor S, Golden TN, Murray A, Abramova E, Malaviya R, Laskin DL, Gow AJ. Myeloid cell dynamics in bleomycin-induced pulmonary injury in mice; effects of anti-TNFalpha antibody. Toxicol Appl Pharmacol. 2021 Apr 15;417:115470. doi: 10.1016/j.taap.2021.115470. Epub 2021 Feb 27.
CrossRefGoogle Scholar
12.Ramos DS, Olivo CR, Quirino Santos Lopes FD, Toledo AC, Martins MA, Lazo Osório RA, Dolhnikoff M, Ribeiro W, Vieira RP. Low-intensity swimming training partially inhibits lipopolysaccharide-induced acute lung injury.Med Sci Sports Exerc. 2010 Jan;42(1):113–9. doi: 10.1249/MSS.0b013e3181ad1c72.
CrossRefPubMedGoogle Scholar
13.Chen Y, Zhou R, Yi Z, Li Y, Fu Y, Zhang Y, Li P, Li X, Pan Y. Porphyromonas gingivalis induced inflammatory responses and promoted apoptosis in lung epithelial cells infected with H1N1 via the Bcl-2/Bax/Caspase-3 signaling pathway. Mol Med Rep. 2018 Jul;18(1):97–104. doi: 10.3892/mmr.2018.8983. Epub 2018 May 7.
CrossRefGoogle Scholar
14.Vieira RP, Müller T, Grimm M, von Gernler V, Vetter B, Dürk T, Cicko S, Ayata CK, Sorichter S, Robaye B, Zeiser R, Ferrari D, Kirschbaum A, Zissel G, Virchow JC, Boeynaems JM, Idzko M. Purinergic receptor type 6 contributes to airway inflammation and remodeling in experimental allergic airway inflammation. Am J Respir Crit Care Med. 2011 Jul 15;184(2):215–23. doi: 10.1164/rccm.201011-1762OC. Epub 2011 Apr 21.
CrossRefPubMedGoogle Scholar
15.Caraccio N, Monzani F, Santini E, Cuccato S, Ferrari D, Callegari MG, Gulinelli S, Pizzirani C, Di Virgilio F, Ferrannini E, Solini A. Extracellular adenosine 5’-triphosphate modulates interleukin-6 production by human thyrocytes through functional purinergic P2 receptors. Endocrinology. 2005 Jul;146(7):3172–8. doi: 10.1210/en.2004-1527. Epub 2005 Mar 24.
CrossRefPubMedWeb of ScienceGoogle Scholar
16.Oliveira CR, Vieira RP. Anti-Inflammatory Activity of Miodesin™: Modulation of Inflammatory Markers and Epigenetic Evidence. Oxid Med Cell Longev. 2020 May 15;2020:6874260. doi: 10.1155/2020/6874260. eCollection 2020.
CrossRefGoogle Scholar
17.Garcia M, Santos-Dias A, Bachi ALL, Oliveira-Junior MC, Andrade-Souza AS, Ferreira SC, Aquino-Junior JCJ, Almeida FM, Rigonato-Oliveira NC, Oliveira APL, Savio LEB, Coutinho-Silva R, Müller T, Idzko M, Siepmann T, Vieira RP. Creatine supplementation impairs airway inflammation in an experimental model of asthma involving P2X7 receptor. Eur J Immunol. 2019 Jun;49(6):928–939. doi: 10.1002/eji.201847657. Epub 2019 Apr 18.
CrossRefGoogle Scholar
18.Mehta P, McAuley DF, Brown M, Sanchez E, Tattersall RS, Manson JJ; HLH Across Speciality Collaboration, UK. COVID-19: consider cytokine storm syndromes and immunosuppression. Lancet. 2020 Mar 28;395(10229):1033–1034. doi: 10.1016/S0140-6736(20)30628-0. Epub 2020 Mar 16.
CrossRefPubMedGoogle Scholar
19.Monteagudo LA, Boothby A, Gertner E. Continuous Intravenous Anakinra Infusion to Calm the Cytokine Storm in Macrophage Activation Syndrome. ACR Open Rheumatol. 2020 May;2(5):276–282. doi: 10.1002/acr2.11135. Epub 2020 Apr 21.
CrossRefGoogle Scholar
20.Ullah MA, Rittchen S, Li J, Hasnain SZ, Phipps S. DP1 prostanoid receptor activation increases the severity of an acute lower respiratory viral infection in mice via TNF-α-induced immunopathology. Mucosal Immunol. 2021 Apr 20:1– 10. doi: 10.1038/s41385-021-00405-7. Epub ahead of print. PMID: 33879829; PMCID: PMC8057290.
CrossRefPubMedGoogle Scholar
21.Stravinskas Durigon T, MacKenzie B, Carneiro Oliveira-Junior M, Santos-Dias A, De Angelis K, Malfitano C, Kelly da Palma R, Moreno Guerra J, Damaceno-Rodrigues NR, Garcia Caldini E, de Almeida FM, Aquino-Santos HC, Rigonato-Oliveira NC, Leal de Oliveira DB, Aimbire F, Ligeiro de Oliveira AP, Franco de Oliveira LV, Durigon EL, Hiemstra PS, Vieira RP. Aerobic Exercise Protects from Pseudomonas aeruginosa-Induced Pneumonia in Elderly Mice. J Innate Immun. 2018;10(4):279–290. doi: 10.1159/000488953. Epub 2018 May 29.
CrossRefPubMedGoogle Scholar
22.Postlethwaite AE, Raghow R, Stricklin GP, Poppleton H, Seyer JM, Kang AH. Modulation of fibroblast functions by interleukin 1: increased steady-state accumulation of type I procollagen messenger RNAs and stimulation of other functions but not chemotaxis by human recombinant interleukin 1 alpha and beta. J Cell Biol. 1988 Feb;106(2):311–8. doi: 10.1083/jcb.106.2.311.
Abstract/FREE Full TextGoogle Scholar
23.Kähäri VM, Heino J, Vuorio E. Interleukin-1 increases collagen production and mRNA levels in cultured skin fibroblasts. Biochim Biophys Acta. 1987 Jul 6;929(2):142–7. doi: 10.1016/0167-4889(87)90169-8.
CrossRefPubMedGoogle Scholar
24.Ambardar SR, Hightower SL, Huprikar NA, Chung KK, Singhal A, Collen JF. Post-COVID-19 Pulmonary Fibrosis: Novel Sequelae of the Current Pandemic. J Clin Med. 2021 Jun 1;10(11):2452. doi: 10.3390/jcm10112452.
CrossRefGoogle Scholar
25.Guimarães LMF, Rossini CVT, Lameu C. Implications of SARS-Cov-2 infection on eNOS and iNOS activity: Consequences for the respiratory and vascular systems. Nitric Oxide. 2021 Jun 1;111-112:64–71. doi: 10.1016/j.niox.2021.04.003. Epub 2021 Apr 6.
CrossRefGoogle Scholar
26.Xu W, Zheng S, Dweik RA, Erzurum SC. Role of epithelial nitric oxide in airway viral infection. Free Radic Biol Med. 2006 Jul 1;41(1):19–28. doi: 10.1016/j.freeradbiomed.2006.01.037. Epub 2006 Feb 20.
CrossRefPubMedGoogle Scholar
27.Kobayashi A, Hashimoto S, Kooguchi K, Kitamura Y, Onodera H, Urata Y, Ashihara T. Expression of inducible nitric oxide synthase and inflammatory cytokines in alveolar macrophages of ARDS following sepsis. Chest. 1998 Jun;113(6):1632–9. doi: 10.1378/chest.113.6.1632.
CrossRefPubMedWeb of ScienceGoogle Scholar
28.Saghir SAM, Al-Gabri NA, Khafaga AF, El-Shaer NH, Alhumaidh KA, Elsadek MF, Ahmed BM, Alkhawtani DM, Abd El-Hack ME. Thymoquinone-PLGA-PVA Nanoparticles Ameliorate Bleomycin-Induced Pulmonary Fibrosis in Rats via Regulation of Inflammatory Cytokines and iNOS Signaling. Animals (Basel). 2019 Nov 11;9(11):951. doi: 10.3390/ani9110951.
CrossRefGoogle Scholar
29.Cinar R, Iyer MR, Kunos G. Dual inhibition of CB(1) receptors and iNOS, as a potential novel approach to the pharmacological management of acute and long COVID-19. Br J Pharmacol. 2021 Mar 26:10.1111/bph.15461. doi: 10.1111/bph.15461. Online ahead of print.
CrossRefGoogle Scholar
30.van de Veerdonk FL, Netea MG. Blocking IL-1 to prevent respiratory failure in COVID-19. Crit Care. 2020 Jul 18;24(1):445. doi: 10.1186/s13054-020-03166-0.
CrossRefGoogle Scholar
31.Franzetti M, Forastieri A, Borsa N, Pandolfo A, Molteni C, Borghesi L, Pontiggia S, Evasi G, Guiotto L, Erba M, Pozzetti U, Ronchetti A, Valsecchi L, Castaldo G, Longoni E, Colombo D, Soncini M, Crespi S, Maggiolini S, Guzzon D, Piconi S. IL-1 Receptor Antagonist Anakinra in the Treatment of COVID-19 Acute Respiratory Distress Syndrome: A Retrospective, Observational Study. J Immunol. 2021 Apr 1;206(7):1569–1575. doi: 10.4049/jimmunol.2001126. Epub 2021 Feb 5.
Abstract/FREE Full TextGoogle Scholar
32.Franzetti M, Pozzetti U, Carugati M, Pandolfo A, Molteni C, Faccioli P, Castaldo G, Longoni E, Ormas V, Iemoli E, Piconi S. Interleukin-1 receptor antagonist anakinra in association with remdesivir in severe COVID-19: A case report. Int J Infect Dis. 2020 Aug;97:215-218. doi: 10.1016/j.ijid.2020.05.050. Epub 2020 May 16.
CrossRefPubMedGoogle Scholar
33.Cavalli G, De Luca G, Campochiaro C, Della-Torre E, Ripa M, Canetti D, Oltolini C, Castiglioni B, Tassan Din C, Boffini N, Tomelleri A, Farina N, Ruggeri A, Rovere-Querini P, Di Lucca G, Martinenghi S, Scotti R, Tresoldi M, Ciceri F, Landoni G, Zangrillo A, Scarpellini P, Dagna L. Interleukin-1 blockade with high-dose anakinra in patients with COVID-19, acute respiratory distress syndrome, and hyperinflammation: a retrospective cohort study. Lancet Rheumatol. 2020 Jun;2(6):e325–e331. doi: 10.1016/S2665-9913(20)30127-2. Epub 2020 May 7.
CrossRefGoogle Scholar
CrossRefPubMedGoogle Scholar
2.de Roquetaillade C, Mansouri S, Brumpt C, Neuwirth M, Voicu S, Le Dorze M, Fontaine C, Barthelemy R, Gayat E, Megarbane B, Mebazaa A, Chousterman BG. Comparison of Circulating Immune Cells Profiles and Kinetics Between Coronavirus Disease 2019 and Bacterial Sepsis. Crit Care Med. 2021 May 18. doi: 10.1097/CCM.0000000000005088. Epub ahead of print. PMID: 34001691.
CrossRefPubMedGoogle Scholar
3.Lange A, Lange J, Jaskuła E. Cytokine Overproduction and Immune System Dysregulation in alloHSCT and COVID-19 Patients. Front Immunol. 2021 Jun 2;12:658896. doi: 10.3389/fimmu.2021.658896. PMID: 34149697; PMCID: PMC8206782.
CrossRefPubMedGoogle Scholar
4.Chua RL, Lukassen S, Trump S, Hennig BP, Wendisch D, Pott F, Debnath O, Thürmann L, Kurth F, Völker MT, Kazmierski J, Timmermann B, Twardziok S, Schneider S, Machleidt F, Müller-Redetzky H, Maier M, Krannich A, Schmidt S, Balzer F, Liebig J, Loske J, Suttorp N, Eils J, Ishaque N, Liebert UG, von Kalle C, Hocke A, Witzenrath M, Goffinet C, Drosten C, Laudi S, Lehmann I, Conrad C, Sander LE, Eils R. COVID-19 severity correlates with airway epithelium-immune cell interactions identified by single-cell analysis. Nat Biotechnol. 2020 Aug;38(8):970–979. doi: 10.1038/s41587-020-0602-4. Epub 2020 Jun 26. PMID: 32591762.
CrossRefPubMedGoogle Scholar
5.Frey A, Lunding LP, Ehlers JC, Weckmann M, Zissler UM, Wegmann M. More Than Just a Barrier: The Immune Functions of the Airway Epithelium in Asthma Pathogenesis. Front Immunol. 2020 Apr 28;11:761. doi: 10.3389/fimmu.2020.00761. eCollection 2020.
CrossRefGoogle Scholar
6.Leyva-Grado VH, Ermler ME, Schotsaert M, Gonzalez MG, Gillespie V, Lim JK, GarcÍa-Sastre A. Contribution of the Purinergic Receptor P2X7 to Development of Lung Immunopathology during Influenza Virus Infection. mBio. 2017 Mar 28;8(2):e00229–17. doi: 10.1128/mBio.00229-17.
CrossRefGoogle Scholar
7.Riteau N, Gasse P, Fauconnier L, Gombault A, Couegnat M, Fick L, Kanellopoulos J, Quesniaux VF, Marchand-Adam S, Crestani B, Ryffel B, Couillin I. Extracellular ATP is a danger signal activating P2X7 receptor in lung inflammation and fibrosis. Am J Respir Crit Care Med. 2010 Sep 15;182(6):774–83. doi: 10.1164/rccm.201003-0359OC. Epub 2010.
CrossRefPubMedWeb of ScienceGoogle Scholar
8.Di Virgilio F, Tang Y, Sarti AC, Rossato M. A rationale for targeting the P2X7 receptor in Coronavirus disease 19. Br J Pharmacol. 2020 Nov;177(21):4990–4994. doi: 10.1111/bph.15138. Epub 2020 Jul 26.
CrossRefGoogle Scholar
9.Chiao CW, Tostes RC, Webb RC. P2X7 receptor activation amplifies lipopolysaccharide-induced vascular hyporeactivity via interleukin-1 beta release. J Pharmacol Exp Ther. 2008 Sep;326(3):864–70. doi: 10.1124/jpet.107.135350. Epub 2008 Jun 16.
Abstract/FREE Full TextGoogle Scholar
10.Sato Y, Chibana K, Horigane Y, Uchida N, Masawa M, Koike R, Nakamura Y, Watanabe T, Shiobara T, Arai R, Shimizu Y, Takemasa A, Ishii Y. Comparison of inducible nitric oxide synthase mRNA expression in different airway portions and association with nitric oxide parameters from patients with asthma. Clin Exp Allergy. 2019 May;49(5):582–590. doi: 10.1111/cea.13344. Epub 2019 Feb 15.
CrossRefGoogle Scholar
11.Venosa A, Gow JG, Taylor S, Golden TN, Murray A, Abramova E, Malaviya R, Laskin DL, Gow AJ. Myeloid cell dynamics in bleomycin-induced pulmonary injury in mice; effects of anti-TNFalpha antibody. Toxicol Appl Pharmacol. 2021 Apr 15;417:115470. doi: 10.1016/j.taap.2021.115470. Epub 2021 Feb 27.
CrossRefGoogle Scholar
12.Ramos DS, Olivo CR, Quirino Santos Lopes FD, Toledo AC, Martins MA, Lazo Osório RA, Dolhnikoff M, Ribeiro W, Vieira RP. Low-intensity swimming training partially inhibits lipopolysaccharide-induced acute lung injury.Med Sci Sports Exerc. 2010 Jan;42(1):113–9. doi: 10.1249/MSS.0b013e3181ad1c72.
CrossRefPubMedGoogle Scholar
13.Chen Y, Zhou R, Yi Z, Li Y, Fu Y, Zhang Y, Li P, Li X, Pan Y. Porphyromonas gingivalis induced inflammatory responses and promoted apoptosis in lung epithelial cells infected with H1N1 via the Bcl-2/Bax/Caspase-3 signaling pathway. Mol Med Rep. 2018 Jul;18(1):97–104. doi: 10.3892/mmr.2018.8983. Epub 2018 May 7.
CrossRefGoogle Scholar
14.Vieira RP, Müller T, Grimm M, von Gernler V, Vetter B, Dürk T, Cicko S, Ayata CK, Sorichter S, Robaye B, Zeiser R, Ferrari D, Kirschbaum A, Zissel G, Virchow JC, Boeynaems JM, Idzko M. Purinergic receptor type 6 contributes to airway inflammation and remodeling in experimental allergic airway inflammation. Am J Respir Crit Care Med. 2011 Jul 15;184(2):215–23. doi: 10.1164/rccm.201011-1762OC. Epub 2011 Apr 21.
CrossRefPubMedGoogle Scholar
15.Caraccio N, Monzani F, Santini E, Cuccato S, Ferrari D, Callegari MG, Gulinelli S, Pizzirani C, Di Virgilio F, Ferrannini E, Solini A. Extracellular adenosine 5’-triphosphate modulates interleukin-6 production by human thyrocytes through functional purinergic P2 receptors. Endocrinology. 2005 Jul;146(7):3172–8. doi: 10.1210/en.2004-1527. Epub 2005 Mar 24.
CrossRefPubMedWeb of ScienceGoogle Scholar
16.Oliveira CR, Vieira RP. Anti-Inflammatory Activity of Miodesin™: Modulation of Inflammatory Markers and Epigenetic Evidence. Oxid Med Cell Longev. 2020 May 15;2020:6874260. doi: 10.1155/2020/6874260. eCollection 2020.
CrossRefGoogle Scholar
17.Garcia M, Santos-Dias A, Bachi ALL, Oliveira-Junior MC, Andrade-Souza AS, Ferreira SC, Aquino-Junior JCJ, Almeida FM, Rigonato-Oliveira NC, Oliveira APL, Savio LEB, Coutinho-Silva R, Müller T, Idzko M, Siepmann T, Vieira RP. Creatine supplementation impairs airway inflammation in an experimental model of asthma involving P2X7 receptor. Eur J Immunol. 2019 Jun;49(6):928–939. doi: 10.1002/eji.201847657. Epub 2019 Apr 18.
CrossRefGoogle Scholar
18.Mehta P, McAuley DF, Brown M, Sanchez E, Tattersall RS, Manson JJ; HLH Across Speciality Collaboration, UK. COVID-19: consider cytokine storm syndromes and immunosuppression. Lancet. 2020 Mar 28;395(10229):1033–1034. doi: 10.1016/S0140-6736(20)30628-0. Epub 2020 Mar 16.
CrossRefPubMedGoogle Scholar
19.Monteagudo LA, Boothby A, Gertner E. Continuous Intravenous Anakinra Infusion to Calm the Cytokine Storm in Macrophage Activation Syndrome. ACR Open Rheumatol. 2020 May;2(5):276–282. doi: 10.1002/acr2.11135. Epub 2020 Apr 21.
CrossRefGoogle Scholar
20.Ullah MA, Rittchen S, Li J, Hasnain SZ, Phipps S. DP1 prostanoid receptor activation increases the severity of an acute lower respiratory viral infection in mice via TNF-α-induced immunopathology. Mucosal Immunol. 2021 Apr 20:1– 10. doi: 10.1038/s41385-021-00405-7. Epub ahead of print. PMID: 33879829; PMCID: PMC8057290.
CrossRefPubMedGoogle Scholar
21.Stravinskas Durigon T, MacKenzie B, Carneiro Oliveira-Junior M, Santos-Dias A, De Angelis K, Malfitano C, Kelly da Palma R, Moreno Guerra J, Damaceno-Rodrigues NR, Garcia Caldini E, de Almeida FM, Aquino-Santos HC, Rigonato-Oliveira NC, Leal de Oliveira DB, Aimbire F, Ligeiro de Oliveira AP, Franco de Oliveira LV, Durigon EL, Hiemstra PS, Vieira RP. Aerobic Exercise Protects from Pseudomonas aeruginosa-Induced Pneumonia in Elderly Mice. J Innate Immun. 2018;10(4):279–290. doi: 10.1159/000488953. Epub 2018 May 29.
CrossRefPubMedGoogle Scholar
22.Postlethwaite AE, Raghow R, Stricklin GP, Poppleton H, Seyer JM, Kang AH. Modulation of fibroblast functions by interleukin 1: increased steady-state accumulation of type I procollagen messenger RNAs and stimulation of other functions but not chemotaxis by human recombinant interleukin 1 alpha and beta. J Cell Biol. 1988 Feb;106(2):311–8. doi: 10.1083/jcb.106.2.311.
Abstract/FREE Full TextGoogle Scholar
23.Kähäri VM, Heino J, Vuorio E. Interleukin-1 increases collagen production and mRNA levels in cultured skin fibroblasts. Biochim Biophys Acta. 1987 Jul 6;929(2):142–7. doi: 10.1016/0167-4889(87)90169-8.
CrossRefPubMedGoogle Scholar
24.Ambardar SR, Hightower SL, Huprikar NA, Chung KK, Singhal A, Collen JF. Post-COVID-19 Pulmonary Fibrosis: Novel Sequelae of the Current Pandemic. J Clin Med. 2021 Jun 1;10(11):2452. doi: 10.3390/jcm10112452.
CrossRefGoogle Scholar
25.Guimarães LMF, Rossini CVT, Lameu C. Implications of SARS-Cov-2 infection on eNOS and iNOS activity: Consequences for the respiratory and vascular systems. Nitric Oxide. 2021 Jun 1;111-112:64–71. doi: 10.1016/j.niox.2021.04.003. Epub 2021 Apr 6.
CrossRefGoogle Scholar
26.Xu W, Zheng S, Dweik RA, Erzurum SC. Role of epithelial nitric oxide in airway viral infection. Free Radic Biol Med. 2006 Jul 1;41(1):19–28. doi: 10.1016/j.freeradbiomed.2006.01.037. Epub 2006 Feb 20.
CrossRefPubMedGoogle Scholar
27.Kobayashi A, Hashimoto S, Kooguchi K, Kitamura Y, Onodera H, Urata Y, Ashihara T. Expression of inducible nitric oxide synthase and inflammatory cytokines in alveolar macrophages of ARDS following sepsis. Chest. 1998 Jun;113(6):1632–9. doi: 10.1378/chest.113.6.1632.
CrossRefPubMedWeb of ScienceGoogle Scholar
28.Saghir SAM, Al-Gabri NA, Khafaga AF, El-Shaer NH, Alhumaidh KA, Elsadek MF, Ahmed BM, Alkhawtani DM, Abd El-Hack ME. Thymoquinone-PLGA-PVA Nanoparticles Ameliorate Bleomycin-Induced Pulmonary Fibrosis in Rats via Regulation of Inflammatory Cytokines and iNOS Signaling. Animals (Basel). 2019 Nov 11;9(11):951. doi: 10.3390/ani9110951.
CrossRefGoogle Scholar
29.Cinar R, Iyer MR, Kunos G. Dual inhibition of CB(1) receptors and iNOS, as a potential novel approach to the pharmacological management of acute and long COVID-19. Br J Pharmacol. 2021 Mar 26:10.1111/bph.15461. doi: 10.1111/bph.15461. Online ahead of print.
CrossRefGoogle Scholar
30.van de Veerdonk FL, Netea MG. Blocking IL-1 to prevent respiratory failure in COVID-19. Crit Care. 2020 Jul 18;24(1):445. doi: 10.1186/s13054-020-03166-0.
CrossRefGoogle Scholar
31.Franzetti M, Forastieri A, Borsa N, Pandolfo A, Molteni C, Borghesi L, Pontiggia S, Evasi G, Guiotto L, Erba M, Pozzetti U, Ronchetti A, Valsecchi L, Castaldo G, Longoni E, Colombo D, Soncini M, Crespi S, Maggiolini S, Guzzon D, Piconi S. IL-1 Receptor Antagonist Anakinra in the Treatment of COVID-19 Acute Respiratory Distress Syndrome: A Retrospective, Observational Study. J Immunol. 2021 Apr 1;206(7):1569–1575. doi: 10.4049/jimmunol.2001126. Epub 2021 Feb 5.
Abstract/FREE Full TextGoogle Scholar
32.Franzetti M, Pozzetti U, Carugati M, Pandolfo A, Molteni C, Faccioli P, Castaldo G, Longoni E, Ormas V, Iemoli E, Piconi S. Interleukin-1 receptor antagonist anakinra in association with remdesivir in severe COVID-19: A case report. Int J Infect Dis. 2020 Aug;97:215-218. doi: 10.1016/j.ijid.2020.05.050. Epub 2020 May 16.
CrossRefPubMedGoogle Scholar
33.Cavalli G, De Luca G, Campochiaro C, Della-Torre E, Ripa M, Canetti D, Oltolini C, Castiglioni B, Tassan Din C, Boffini N, Tomelleri A, Farina N, Ruggeri A, Rovere-Querini P, Di Lucca G, Martinenghi S, Scotti R, Tresoldi M, Ciceri F, Landoni G, Zangrillo A, Scarpellini P, Dagna L. Interleukin-1 blockade with high-dose anakinra in patients with COVID-19, acute respiratory distress syndrome, and hyperinflammation: a retrospective cohort study. Lancet Rheumatol. 2020 Jun;2(6):e325–e331. doi: 10.1016/S2665-9913(20)30127-2. Epub 2020 May 7.
CrossRefGoogle Scholar
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





Development and research of the company "LibiPharm" - "LibiPharm" Israel.
© 2023 Calpton SIA. All rights reserved.
We accept
